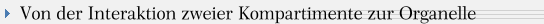
 Die Endosymbionten-Hypothese der Biologie
besagt, dass eine Reihe von Zellorganellen in den Zellen von Eukaryoten durch Einverleiben oder Fusion
von unterschiedlichen Organismen entstanden seien (A.F.W. Schimper 1883,
K. Mereschkowski 1905, L. Margulis 1967). Es ließ sich vielfach nachweisen, dass manche Bakterien wie Wolbachia (s. Abb.) ihre Gene nicht nur auf andere Prokaryoten, sondern großzügig auf sogen. höhere Organismen übertragen. Dies hatte auch zu einigen Fehlinterpretationen von Phänomenen bei Mutationsexperimenten an Drosophila geführt.
Die Endosymbionten-Hypothese der Biologie
besagt, dass eine Reihe von Zellorganellen in den Zellen von Eukaryoten durch Einverleiben oder Fusion
von unterschiedlichen Organismen entstanden seien (A.F.W. Schimper 1883,
K. Mereschkowski 1905, L. Margulis 1967). Es ließ sich vielfach nachweisen, dass manche Bakterien wie Wolbachia (s. Abb.) ihre Gene nicht nur auf andere Prokaryoten, sondern großzügig auf sogen. höhere Organismen übertragen. Dies hatte auch zu einigen Fehlinterpretationen von Phänomenen bei Mutationsexperimenten an Drosophila geführt.
Vertikal übertragene Endosymbionten sind in ihrer
Reproduktion vom Überleben und der Fortpflanzung des Wirts abhängig. Dies
begünstigt die Evolution von Kooperation, wie das Beispiel der Mitochondrien
und ihrer eukaryotischen Wirtszellen eindrucksvoll belegt.
Der Evolution dieser
Kooperation können aber auch Hindernisse im Weg stehen, die in der Biologie
zunächst nur theoretisch erkannt wurden. Endosymbionten mit ausschließlich
cytoplasmatischer Vererbung werden von männlichen Wirten nicht auf die nächste
Generation übertragen. Sie stehen daher unter "Selektionsdruck", das
Geschlechtsverhältnis ihrer Wirte zugunsten von Töchtern zu beeinflussen und
sich generell eher an weibliche als an männliche Aspekte der Physiologie ihrer
Wirte anzupassen.
Intrazelluläre Bakterien der Gattung Wolbachia stellen
heute das Modellsystem dar, an dem diese Hypothese besonders eingehend
untersucht wird. Wolbachien gehören phylogenetisch zu den nächsten Verwandten
der Mitochondrien. Sie werden im Regelfall cytoplasmatisch vererbt und greifen
tatsächlich bei vielen Arthropoden massiv in deren Reproduktions- und
Entwicklungsphysiologie ein (Arnold, Berlin).
So sind Plastiden nach der heute als bewiesen geltenden
Endosymbiontentheorie aus den ursprünglich frei lebenden Cyanobakterien
entstanden, die von einem heterotrophen, wohl begeißelten, eukaryonten Wirt
aufgenommen wurden. Der "Erfolg" der primären Endosymbiose stellt sich auch
hier wieder als Gentransfer vom Endosymbionten zur Wirtszelle dar und in der
Fähigkeit, die Produkte der nunmehr eukaryotisch mutierten ehemaligen
Cyanobakteriengene wieder in den Endosymbionten reimportieren zu können.
Der
Verlust genetischer Information ließ den primären Endosymbionten zum Organell
mit prokaryontem Zellwandrest werden (cytoplasmatische Plastidgenese).
Plastiden mit mehr als zwei Hüllmembranen sind das Ergebnis
sekundärer Endosymbiosen: Darunter versteht man die Aufnahme eines
photosynthetischen Eukaryoten durch einen heterotrophen Wirt, also eine
Eukaryoten-Endosymbiose. Während der Umwandlung sekundärer Endosymbiosen in
genetisch stabile komplexe Zellen ist, wie bei der primären Endosymbiose,
Gentransfer und vor allem differentieller Genverlust das entscheidende Phänomen
für den Komplexitätsgrad einer Zelle gewesen (cf. Reduktion von genetischer
"Redundanz"; Kowallik, Düsseldorf). Neuzeit-Beispiel: Synchroma grande, eine Meeresalge in Symbiose mit Paketen von verschachtelten Chloroplasten (= Cyanobakteriennachfahren) nach Schnetter & Wilhelm, 2007.
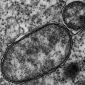
 Cyanobacteria von Australian Bitter Springs Chert (Alter
850 Mio. Jahre) : eine coloniale chroococcalean Form, wahrscheinlich Myxococcoides
minor - Ursprungsform der Komplexbausteine von Eukaryoten. (s. Abbildung) Cyanobacteria von Australian Bitter Springs Chert (Alter
850 Mio. Jahre) : eine coloniale chroococcalean Form, wahrscheinlich Myxococcoides
minor - Ursprungsform der Komplexbausteine von Eukaryoten. (s. Abbildung)
Der als Kometengestein identifizierte "Orgueil-Meteorit" (1864 Impact in Frankreich) ist kürzlich wieder als kosmischer Träger von Strukturen chroococcaleanen Aussehens von der NASA ins Gespräch gebracht worden. REM-Untersuchungen weisen eher auf Magnetite hin. Unbenommen bleibt die Hypothese von Wasserzufuhr durch wiederkehrende Kometen-, Asteroid- und Impactprodukt-Körper beim Aufprall auf die Erde. Grundlagen zu dieser Annahme schufen Newton, Halley et al. (Theorie einer Asteroidenkollision vor 65 Mio. Jahren mit Deep Impact auf der Erde, nach W.Bottke, D.Nesvorny & D.Vokrouhlicky, 2007).
|


